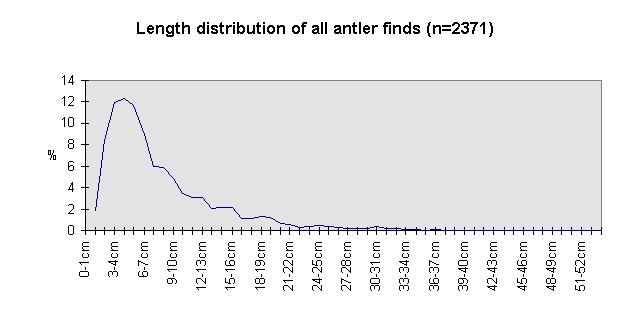
Fig. 6 (left): Length of all antler finds.
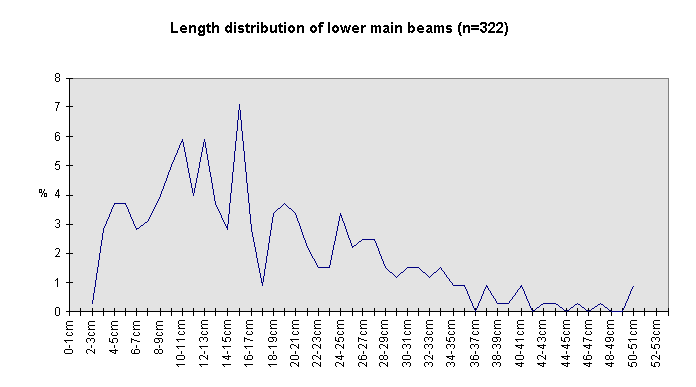
Fig. 7 (right): Length of all lower beams.
An estimate of the minimum number of heads cannot be based on the shed antler material alone. The 193 finds of shed antlers only indicate that a number of male cervids shed their antlers around Bilzingsleben during the Middle Pleistocene, whereas the 157 finds of unshed antlers definitely represent animals which died. Disregarding the cause of death of these cervids, it is possible to express the minimum number of heads represented by their unshed antlers. 14 reconstructed unshed antlers represent 7 animals. All other unshed antler fragments have to be interpreted as singular occurrences, with every piece representing a different animal, totalling 150 animals on the basis of unshed antlers.
Although a preliminary count of the most frequently encountered non-antler cervid material (molars and astragali) suggests the presence of some 80 animals, neither taxonomic nor sex differences were taken into account. It is probable that after excluding females and fallow deer, heads of male red deer were far fewer than 80. It can be expected however that the unshed antlers alone (150 heads) represent more than double the number of male red deer as represented by finds other than antler.
The disproportionate representation of unshed antlers versus other cranial and postcranial skeletal elements of cervids could be viewed as an argument for the selective treatment of antlers by hominids (e.g. collection). On the other hand, the suggestion that parts of the Bilzingsleben faunal material, including antlers, form a naturally deposited assemblage cannot be ignored. Interpretation cannot be made on the strength of antlers alone; comparative results from further taphonomic studies of other material remain to be seen.
To estimate the age of a deer on the basis of an antler must be regarded as uncertain, much less certain than using teeth for example. For unshed antlers rough age estimates are best calculated by the ratio of pedicle length to pedicle diameter (where roughly the greater the value, the older the animal); for shed antlers the 'Formzahl Siegel/Rose' shows some correlation with the age of a red deer (Habermehl 1985). Comparing the coefficients of shed antlers and unshed antlers with values for red deer of recent times (Hattemer and Drechsler 1976; Habermehl 1985), one could deduce that shed antlers at Bilzingsleben mainly derived from juvenile and young adult animals, while unshed antlers mainly came from young adult and adult animals.
Assuming that the rhythm of insolation (exposure to sun rays) during the Middle Pleistocene was similar to that of today, antlers would have been shed in late winter and early spring. Of more taphonomic relevance is the observation that very few of the unshed antlers have demarcation lines in the distal pedicle area (Bubenik 1966). Demarcation lines develop during or slightly after the rut (when deer are physiologically and behaviorally capable of reproduction), hence in autumn, and it is along those lines that annual antler loss occurs. For nearly all unshed antlers, the time of the red deers' death can be narrowed to late summer / early autumn i.e. before the rut.
Evaluation of age structure and aspects of seasonality of the Bilzingsleben cervids must wait until results from the analysis of cranial (non antler) and postcranial skeletal parts are available. Before attempting a meaningful interpretation of such data, how the finds accumulated must also be assessed - such investigations have only just started at Bilzingsleben.
The size class distribution of most of the antler material (Fig. 6) from the 1969-1993 excavations is dominated by small fragments (2591 finds were measured). Lower beam fragments (Fig. 7) and crown fragments (Fig. 8) have a wider size range and a less regular size distribution than isolated tines and fragments of antler cortex of less than 150 mm (Fig. 9).
Fig. 6 (left): Length of all antler finds.
Fig. 7 (right): Length of all lower beams.
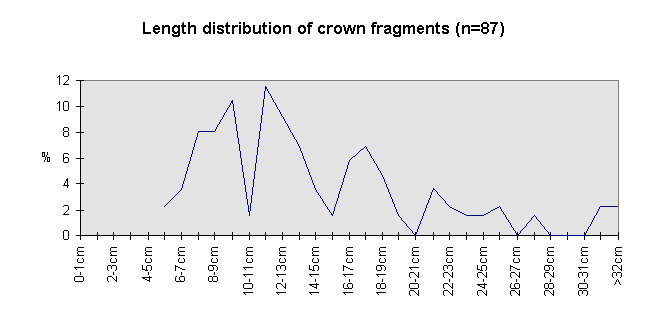
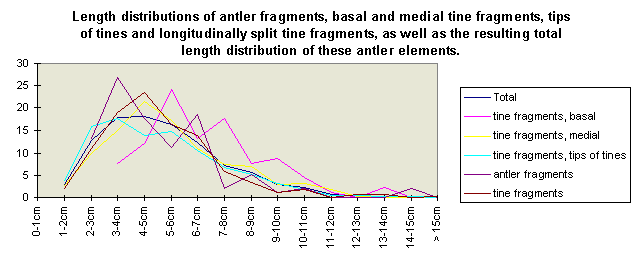
Fig. 8 (left): Length of crown and crown elements.
Fig. 9 (right): Length of tine and antler fragments.
Most of the smaller elements range from 30-60 mm. A total lack of antler parts measuring less than 30 mm has to be investigated, since experiments showed (Fig. 10) that antler artefact production brings about a higher amount of fragments of less than 30 mm than of the 30-60 mm range.
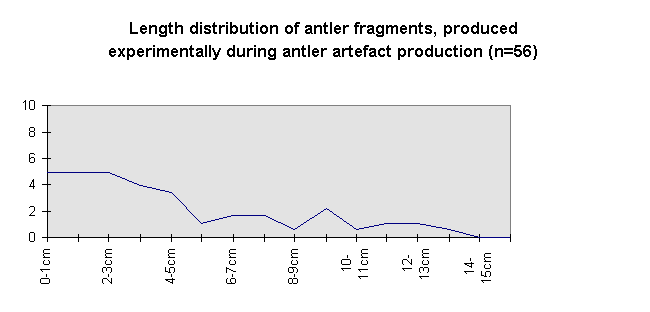
Fig. 10: Length of antler fragments from experimental fragmentation.
Data for the systematic comparison of antler size distribution with those of other faunal material are still not available. Summaries of length distributions for a variety of bone tools and of several groups of stone artefacts are given by Mania (1993b table 2; for lithic artefact dimensions see also Weber 1986).
A few antler fragments are heavily crushed by the weight of sediment overburden, although nearly all of them retain their original shape and are only rarely slightly deformed. The antler material suffers, however, as often from weathering cracks as from abrasion and chemical corrosion. Besides these dominant phenomena, slight manganese staining and modern damage are often present on the material. Forty-seven pieces of antler (1.67% of the total material) exhibit traces of gnawing, most of which can be related to gnawing by other cervids (L. Maul, Weimar (pers. comm) confirms that none of these finds exhibit traces of rodent gnawing). Three pieces of antler show partly eroded traces of root etching. Scratches are abundant, and can in part be related to cervid activities during their lifetime. The Bilzingsleben antler material displays three stages of general surface preservation:
In general, therefore, the surface preservation can be classified as good. Of further interest are observations of differential preservation. All Stage 2 and Stage 3 finds show traces of corrosion. 39.5% of these finds have differentially corroded surfaces, the more corroded surface facing downward when excavated. The remaining 60.5% are evenly corroded on all surfaces. Corrosion is interpreted as a diagenetic phenomenon, probably resulting from the circulation of 'aggressive' ground water (D. Mania, pers. comm.). 42.2% of the Stage 2 and Stage 3 finds show a greater degree of abrasion on one of their faces, while 57.8% are evenly abraded. Finds showing greater abrasion on one side were found with that side facing upward.
Varying abrasion occurs in all areas of the excavated landscape and, as far as it is documented, from all stratigraphic units. The same is true for bilaterally abraded finds. The abrasive forces of running surface water have a different effect on antlers and other bones, resulting in the re-deposition of the majority of antler finds before final burial, followed by another phase of fluvial abrasion.
© Internet Archaeology
URL: http://intarch.ac.uk/journal/issue8/vollbrecht/en/5.html
Last updated: Thu Aug 10 2000